 |
| Figur 1 |
MEDVETANDE SOM SIMULERING AV BETEENDE OCH PERCEPTION
Germund Hesslow
Institutionen för Fysiologiska vetenskaper
Lunds universitet
Inledning
"Medvetandeproblemet", som tills ganska nyligen av många ansågs vara ett utomvetenskapligt problem, har under senare år blivit högsta mode. Många inom hjärnforskningens jetset deltar nu i en kapplöpning där evig ära och berömmelse väntar den som presenterar den slutgiltiga lösningen på problemet. Man kan emellertid hävda - det tänker jag i alla fall göra här - att delar av problemet redan har lösts eller åtminstone att det gradvis håller på att lösas upp inför våra ögon. De viktigaste av de idéer och empiriska resultat som krävs för att förstå medvetandet finns redan. Några av dem har i själva verket funnits i mer än ett sekel.
Låt oss först konstatera att det, som ibland kallas medvetandeproblemet, i själva verket utgörs av en lång rad av sammanlänkade frågor. Det kan vara instruktivt att begrunda ett annat klassiskt biologiskt problem, nämligen frågan vad "liv" egentligen är. För bara några decennier sedan betraktades detta som ett stort mysterium och debatterades passionerat av vetenskapsmän och filosofer. Vitalismen betraktades som en seriös ståndpunkt och Jacques Monod (1970) lyckades uppröra tusentals läsare genom att påstå att livet kunde förklaras i biokemiska termer. Vi bekymrar oss inte längre över detta, inte därför att någon till sist kom på den korrekta lösningen, utan därför att problemet har ersatts av en rad mera specifika frågor, som "Hur lagrar en cell energi?", "Hur reproducerar den sig?" eller "Hur håller den sin interna kemiska miljö konstant?", för vilka det nu finns specifika svar. Det finns skäl att tro att också medvetandeproblemet kommer att lösas upp i ett antal separata frågor. För många hjärnforskare, såväl som lekmän, torde följande frågor tillhöra de mest centrala:
Hur uppkommer den inre världen? För de flesta människor är antagligen detta den mest centrala frågan. Hur kan det komma sig att jag kan stänga mina ögon och ändå se föremål som saknar fysisk realitet? Ett för många hjärnforskare besläktat problem är att det inte tycks finnas någon struktur i hjärnan som är avdelad för medvetande eller tänkande. Det finns sensoriska och motoriska områden i hjärnan men inga specifikt kognitiva och de ännu inte kartlagda områden som blir över för medvetet tänkande blir allt mindre. Dessutom tyder nya resultat på att vissa områden som av tradition betraktats som renodlat motoriska som cerebellum och de basala ganglierna också deltar i kognitiva processer.
Vad är mentala objekt? Vad är det jag ser när jag ser något för mitt inre öga? Om jag tänker på, låt oss säga, en enhörning, tänker jag på något som saknar materiell existens, men vad består det av om det inte består av materia? Mental substans?
Vad är en tanke? Medan känslor, önskningar och drifter av somliga anses kunna vara omedvetna antas tänkande nästan alltid vara en medveten process och intimt förknippat med medvetandet. Att tänka på något är i det närmaste synonymt med att vara medveten om det. Att förstå vad en tanke är för någonting är därför en viktig del av medvetandets problem.
Funktionsproblemet. Varför finns den inre världen? Borde inte evolutionen ha kunnat skapa en organism som på vissa givna stimuli reagerar med samma beteende som en människa men utan att detta medieras av en inre värld? Sådana zombies borde var omöjliga att skilja från människor och de borde kunna överleva och reproducera sig lika bra som människor. Så varför har inte evolutionen nöjt sig med zombies? Vilken biologisk fördel har den inre världen?
Vilken sorts varelser kan ha en inre värld? Kan djur ha
en inre värld? Robotar?
Jag skall här beskriva en fysiologisk mekanism med vars hjälp dessa frågor kan besvaras. Den centrala idén är att hjärnan kan generera sitt eget inflöde. Mekanismen har tre komponenter:
1) Simulering av beteende. Att tänka att man gör något är nästan samma sak som att faktiskt göra det, men handlingen fullbordas inte.
2) Simulering av perception. Att föreställa sig att man ser (hör, känner) något är nästan samma sak som att faktiskt se (höra, känna) något, men den perceptuella aktiviteten är genererad av hjärnan själv i stället för av utifrån kommande stimuli.
3) Antecipation. Vi antar också associationsmekanismer
som gör det möjligt för simulering av såväl beteende
som perception att utlösa aktivitet i hjärnans sensoriska områden.
En konsekvens av detta är att simulering av beteende kan utlösa
perceptuell aktivitet som liknar den aktivitet som sannolikt skulle ha
uppkommit om de simulerade rörelserna faktiskt hade utförts.
Simulering av beteende.
De tidiga behavioristerna ansåg att tänkande inte var något annat än vad de kallade "förberedande", "incipient" eller "dolt" beteende ("covert behavior"), dvs. rörelser i extremiteter eller talmuskulatur som utförs så svagt att de inte ger upphov till iakttagbara rörelser men som ändå kan registreras med elektrofysiologiska tekniker. En liknande idé hade tidigare formulerats av Alexander Bain (1868). "The tendency of the idea of an action to produce the fact, shows that the idea is already the fact in a weaker form. Thinking is restrained speaking or acting" (s. 340). Dessa svaga rörelser, tänkte man sig, var tillräckliga för att aktivera led- och muskelsinnena vilket ger upphov till stimuli som kan utlösa nya svaga muskelrörelser. Varje länk i denna kedja av stimuli och responser tänktes vara inlärda s.k. betingade reflexer (Watson, 1930). Många fysiologer lade ner stora ansträngningar på att visa att tankar faktiskt brukar åtföljas av svag elektrisk aktivitet muskulaturen (Jacobson, 1930; Sokolov, 1972). Hypotesen kom dock i vanrykte när det visade sig att personer vars muskler fullständigt förlamats av curarepreparat ändå var fullt kapabla till normalt tänkande (Smith et al., 1947). Dessutom påpekade Lashley (1951) att sådana reflexkedjor skulle vara alldeles för långsamma för att kunna förklara tänkandet. Det tar helt enkelt för lång tid för den nervsignal som först måste gå från hjärnan ut till muskeln och sedan hela vägen tillbaka till hjärnan innan nästa länk i kedjan kan utlösas. Behavioristernas grundläggande idé, att en tanke är ett förstadium till det fullbordade beteendet, kan emellertid ges en något modifierad tolkning och har faktiskt vunnit anklang och empiriskt stöd i modern neurofysiologi.
Komplexa handlingar kan beskrivas som hierarkiskt organiserade komponenter,
så att ett övergripande mål delas upp i delmål och
delhandlingar ända ner till den enskilda muskelcellen. Handlingen
att laga middag består av komponenter som att ta fram ingredienserna,
stycka dem, koka och steka, lägga upp, duka mm. Dessa komponenter
kan i sin tur delas upp i andra. Att stycka en bit kött består
av ganska komplicerade kombinationer av rörelser, vilka i sin tur
består av mer elementära rörelser och så vidare ner
till de enkla muskelsammandragningarna. Det förekommer att man beskriver
språkligt beteende, som ju uppenbart kan beskrivas som en hierarkisk
struktur, som om språket vore unikt i detta avseende (Pinker, 1994)
men nästan allt intressant beteende har i själva verket en sådan
organisation.
|
| Figur 1 |
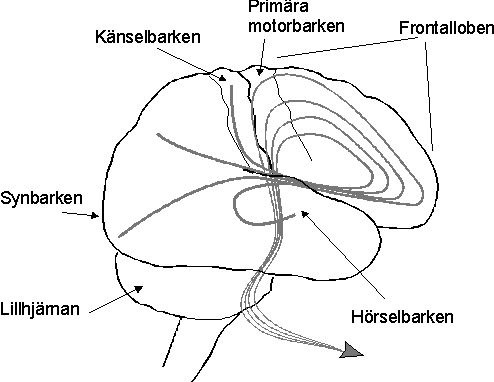
Mänskligt beteende genereras i hjärnans frontallober som svar på sensoriskt inflöde (se figur 1). Externa stimuli aktiverar primära sensoriska områden, vilka i sin tur aktiverar vad man ibland kallar sensoriska associationsområden. Ju längre man kommer från det primära området desto mer subtila och abstrakta är de egenskaper hos stimuli som aktiverar nervcellerna. En grupp celler i den primära synbarken reagerar t.ex. på belysning med en viss intensitet av en specifik punkt på näthinnan. Denna information säger dock inte i sig särskilt mycket om det observerade föremålets egenskaper. En starkt belyst svart yta kan reflektera mer ljus än en ljus yta i mörker. För att avgöra hur det observerade föremålet självt är beskaffat måste belysningen av näthinnan kombineras med annan information såsom belysningsförhållanden och hur slät ytan är. Information om var på näthinnan ett objekt projiceras säger något om var objektet befinner sig i verkligheten bara i kombination med information om huvudets läge och blickens riktning. Sådana mer abstrakta aspekter som en ytas reflekterande egenskaper eller lokalisationen av ett föremål motsvaras av aktivitet i nervceller i visuella associationsområden.
De sensoriska signalerna går sedan vidare till frontalloberna för att generera ett för situationen adekvat beteende. I det område som kallas den primära motorbarken ligger nervceller som signalerar till enskilda muskler eller muskelgrupper, exempelvis att ett finger skall böjas eller en hand lyftas. Man kan säga att dessa nervceller svarar för de beteendekomponenter som befinner sig längst ner i en hierarkiskt organiserad handling. För att generera en mera komplex rörelse, högre upp i hierarkin, som att gripa eller lyfta ett föremål, måste flera muskler, och således flera cellgrupper i primära motorbarken, aktiveras i en bestämd sekvens. Denna måste också ta hänsyn till en mängd tillfälliga situationsbetingade omständigheter, som kroppens och handens aktuella positioner och föremålets form och vikt. Signalerna för att göra detta kommer från områden längre fram i frontalloberna med bistånd från lillhjärnan. Ännu mer komplexa beteenden som innefattar sekvenser av handlingar, styrs från områden ännu längre fram. Enkelt uttryckt kan man säga att ju längre fram man går, desto mer globala aspekter på beteendet styrs därifrån och desto högre upp i hierarkin kommer man.
När jag i det följande talar om att man kan simulera en handling menar jag helt enkelt att man kan aktivera frontalloberna på ungefär samma sätt som när man utför en fysiskt fullbordad handling, men att signalflödet bromsas strax innan de motoriska signalerna lämnar hjärnan. En simulerad handling är således ungefär detsamma som en fysiskt ofullbordad handling. Det bör kanske understrykas att frontallobsaktiviteten vid en simulerad handling inte kan vara exakt likadan som vid en fullbordad handling, ty i så fall skulle denna aktivitet rimligen ha samma effekter, dvs leda till en fullbordad handling. Det hypotesen kräver är en hög grad av likhet, att en simulerad och en fullbordad handling aktiverar i huvudsak samma områden och samma nervceller och att mekanismerna i huvudsak är desamma. Det finns en omfattande vetenskaplig litteratur som stöder denna hypotes (se Jeannerod, 1994 för en översikt) och här skall jag endast kortfattat nämna några exempel.
Beteendeförsök har visat flera paralleller mellan simulerade och faktiskt utförda rörelser. Det har t.ex. länge varit känt inom idrottsrörelsen att en mental simulering förbättrar senare prestationer, vilket ju talar för att simuleringen påverkat vissa av de nervkretsar som senare används vid den verkliga idrottsliga uppgiften. Flera studier visar att den tid det tar att simulera en motorisk uppgift motsvarar den tid det tar att faktiskt utföra samma uppgift. Att t.ex. läsa upp alfabetet tyst för sig själv tar ungefär samma tid som att göra det högt (Landauer, 1962). I ett annat experiment fick försökspersoner gå med förbundna ögon till olika platser som de tidigare fått se, samtidigt som de med ett stoppur fick indikera när de började och när de ansåg sig ha nått målet. När de sedan fick föreställa sig att de gick till samma platser, visade det sig att det förelåg en mycket god överensstämmelse mellan den tid en viss person tog på sig för den inbillade respektive den faktiska promenaden (Decety et al., 1989).
Mera direkta belägg för simuleringshypotesen kommer från mätningar av regionalt blodflöde. Ökad aktivitet i ett visst hjärnområde leder till ökad ämnesomsättning, vilket i sin tur ger ett ökat blodflöde, vilket kan mätas med olika tekniker. Ett stort antal blodflödesstudier visar att försökspersoner som instrueras att föreställa sig vissa rörelser aktiverar delvis samma delar av hjärnan som aktiveras när man faktiskt utför dessa rörelser. Ingvar och Philipsson (1977) visade att både försökspersoner som instruerades att mentalt simulera respektive faktiskt utföra handrörelser fick en ökad aktivitet i frontallobernas främre delar, medan bara den andra gruppen aktiverade den motoriska hjärnbarken. En lång rad senare studier har bekräftat dessa fynd. Överensstämmelsen är inte helt perfekt. Ett område som kallas den supplementära motorarean aktiveras t.ex. vid både simulerad och utförd rörelse men det tycks vara delvis olika delar som aktiveras (Deiber et al., 1998).
Man har ganska nyligen påvisat en ökad excitabilitet i den motoriska hjärnbarken när försökpersoner simulerar vissa rörelser. Man kan aktivera relativt små områden i hjärnans motoriska bark med hjälp av en teknik som kallas "transkraniell magnetisk stimulering". Om man stimulerar barken på ett sådan sätt, att man aktiverar en viss muskelgrupp före respektive efter man bett försökspersonen att tänka på samma rörelse, kommer stimuleringen att utlösa en kraftigare rörelse i det andra fallet (Bonnet et al., 1997). Detta tyder på att tanken, att man utför en viss rörelse, stimulerar de nervceller i den motoriska barken som normalt utför rörelsen.
Det kan i detta sammanhang även vara värt att nämna upptäckten
av s.k. "spegelneuron", ett fynd som väckt stor uppmärksamhet
under senare år. Om man registrerar aktiviteten hos enskilda nervceller
i frontalloben på en apa, så finner man celler som aktiveras
specifikt när apan utför vissa rörelser. Någon cell
kan aktiveras när apan lyfter handen, en annan när den går
baklänges. Det har nu visat sig i flera försök att vissa
av dessa nervceller (spegelneuronen) också kan aktiveras när
apan passivt iakttar en annan apa eller en människa som utför
samma rörelse (Di Pellegrino et al., 1992). Vilken funktion
dessa nervceller har är omdiskuterat, men om man utifrån hypotesen
att tanken att man utför en rörelse i hög grad är samma
sak som förberedelsen för motsvarande fullbordade rörelse,
är förekomsten av dessa spegelneuron inte så förvånande.
Om en apa alls kan tänka på att lyfta handen, borde ett ganska
pålitligt sätt att inducera denna tanke vara att visa apan någon
annan som lyfter handen. När den tänker på en rörelse,
utför den en simulering, vilket innebär aktivering av många
av de nervceller som normalt aktiveras vid motsvarande fysiskt fullbordade
rörelse.
Simulering av perception
Om jag skär mig i fingret, kommer nervimpulser från den skadade huden att skicka impulser till hjärnan där de, i hög grad okända, processer äger rum, som vi kallar perceptionen och upplevelsen av skadan. Om de perifera nervfibrerna kunde aktiveras på artificiell väg, någonstans på vägen från fingret till hjärnan, skulle naturligtvis samma upplevelse uppkomma. Det är något i denna stil som ibland händer patienter som fått en kroppsdel amputerad. De skadade nervfibrerna från t.ex. en hand kan fortsätta att skicka impulser till hjärnan och ge upphov till förnimmelser av den icke existerande kroppsdelen. Patienten sägs ha en "fantomlem", ibland även s.k. "fantomsmärta". Exemplet visar att en upplevelse av något inte måste betyda att det existerar något som upplevs. Vi behöver inte anta att den fingerskada som kan upplevas från den amputerade handen innebär att det existerar en fiktiv hand eller en fiktiv skada.
Om man applicerar ett lätt tryck på ögongloben kan man ibland "se" stjärnor eller blixtar inte därför att man egentligen tittar på mentala stjärnor och blixtar, utan därför att man simulerar den aktivitet i synbanorna som normalt initieras av faktiska stjärnor och blixtar. Detsamma borde gälla om vi kunde aktivera hjärnans synområden på ett sätt som liknar den aktivitet som uppkommer när man iakttar ett materiellt objekt. Om jag kunde simulera den aktivitet som uppkommer när jag tittar på en bil, borde jag också "se" en bil. Eftersom den process i hjärnan som normalt pågår när vi ser en bil är orsakad av en bil, är det frestande att tänka sig att denna process i frånvaro av en fysisk bil måste orsakas av en annan sorts bil, en som är icke-fysisk eller mental. Men detta är, som vi nu bör kunna inse, felaktigt.
Det är viktigt att notera att vi inte har gjort något antagande här om vari den perceptuella aktiviteten egentligen består. I synnerhet har vi inte antagit att hjärnan konstruerar någon sorts kopia, bild eller "representation" av verkligheten. Sensoriskt inflöde leds, via komplicerade nervösa vägar som vi inte förstår så mycket av, till beteende. Enkla versioner av sådana händelsekedjor förekommer i primitiva organismer och kan enkelt implementeras i tekniska konstruktioner som robotar eller enkla leksaker. I min bil leds en signal från hjulen till hastighetsmätaren som kan sägas "observera" bilens hastighet. Det är såväl onödigt som vilseledande att säga att systemet har konstruerat en "bild" eller en "representation" av hastigheten.
Om ett mekaniskt föremål har konstruerats för att reagera på ett viss sätt på en viss signal, är det i allmänhet möjligt att kringgå den normala insignalen och, så att säga, skjuta in en "falsk" signal före den mekanism som genererar reaktionen. En falsk signal skulle t.ex. kunna få min hastighetsmätare att visa en viss hastighet även när bilen står stilla. Detta gäller naturligtvis också om reaktionen råkar vara en verbal beskrivning av insignalen. Min dator kan observera (utan citattecken) vilka tangenter som just har tryckts ned och ett enkelt program kan skriva på skärmen "jag känner att du just tryckte på Y-tangenten". Det vore inte svårt att lura programmet att skriva detta genom att skicka en falsk Y-signal från tangentbordet till datorn utan att faktiskt ha tryckt på Y-tangenten. Ur datorns synvinkel vore denna signal oskiljaktig från den som normalt signalerar att Y har trycks ned. Vi kan säga att datorn simulerar observationen att Y just tryckts ned. I detta fall kan man invända att termen "simulering" normalt antyder att det finns en agent som har en viss avsikt med simuleringen. Här använder vi emellertid termen även om den rent passiva processen att initiera en händelsekedja via något annat än det som normalt initierar den.
Jag har än så länge antagit, utan att anföra några belägg för saken, att vi faktiskt kan simulera perception genom att aktivera hjärnans sensoriska områden på ett sätt som liknar aktiviteten vid perception av yttervärlden. Detta är en gammal tanke, som förebådas av David Hume och som explicit formulerats av flera författare på 1800-talet. William James (1890) skriver om den som allmänt accepterad. "The commonly-received idea [about imagination] is that it is only a milder degree of the same process which took place when the thing now imagined was sensibly perceived" (p 68) och han citerar Bain (1868) "the mode of existence of a sensation existing after the fact is essentially the same as its mode of existence during the fact". Denna tanke framfördes utan några egentliga empiriska data som grund, men det finns idag ett mycket omfattande stöd för den både från kognitiv psykologi och från neurovetenskap (se t.ex. Farah, 1988 och särskilt Kosslyn, 1994). Här må det räcka att nämna några exempel.
I en lång rad experiment har Shepard och medarbetare jämfört effekter av konkret manipulering av objekt med motsvarande "mentala" manipulationer. I ett klassiskt experiment fick försökspersoner se teckningar av tredimensionella föremål bestående av fyrkantiga boxar sammansatta på olika sätt. Uppgiften att avgöra om två föremål var identiska skulle kunnat lösas genom att fysiskt rotera den ena tills den, eventuellt, sammanfaller med den andra, men försökspersonerna måste göra detta "i huvudet". Intressant nog visar det sig att den tid det tar att lösa uppgiften samvarierar starkt med hur mycket objektet måste roteras för att sammanfalla med referensobjektet, ungefär som om man betraktade ett roterande objekt och var tvungen att invänta den position där objekten kan jämföras (Shepard & Metzler, 1971).
Detta och andra liknande experiment har tolkats som belägg för att fantasiföreställningar har "visuella egenskaper" och därför sannolikt har delvis samma mekanismer som synsystemet (Kosslyn, 1994). Dessa experiment är svåra att värdera och alternativa tolkningar har framförts av många forskare (Pylyshyn, 1984). Experiment av dessa slag har också tolkats som belägg för att föreställningar har "bildlika" egenskaper. Denna idé sammanblandas ofta med, men är helt skild från, frågan om fantasiföreställningars neuronala underlag.
En annan typ av belägg kommer från patienter med skador i sensoriska barkområden. Fastän symptomen av sådana skador inte är alldeles entydiga, finns det många exempel på patienter som drabbats av s.k. kortikal blindhet p.g.a. skador i synbarken och som också tappar förmågan att skapa visuella föreställningar (Farah, 1988).
Ett annat slående exempel är de skador i den ena hjärnhalvans hjässlob som leder, inte till blindhet, men till en oförmåga att lägga märke till och reagera på ting i den motsatta delen av synfältet. Detta fenomen, som brukar kallas "neglect", har också en kognitiv parallell. Patienter med vänstersidig "neglect" p.g.a. högersidiga hjässlobsskador ombads föreställa sig att de stod vid ena änden av det berömda torget Piazza del Duomo i sin hemstad Milano och beskriva de byggnader och andra objekt de kunde erinra sig. Patienterna kunde bara beskriva objekt på torgets högra sida i förhållande till den tänkta utsiktspunkten. När de sedan ombads föreställa sig att de stod på torgets motsatta sidan, kunde de beskriva de föremål som nu befann sig i det högra synfältet men som de inte kunde erinra sig i den första omgången (Bisiach & Luzzatti, 1978).
Det viktigaste stödet för idén att visuella föreställningar utnyttjar samma mekanismer som synen har uppnåtts vid försök att direkt mäta aktiviteten i hjärnan när försökspersoner föreställer sig att de ser någonting. I ett tidigt sådant experiment mätte man den elektriska aktiviteten i hjärnans syn- respektive känselområden samtidigt som försökspersonerna fick föreställa sig olika visuella respektive taktila stimuli. När uppgiften var att föreställa sig ljusblixtar ökade nervcellsaktiviteten specifikt i synbarken. När försökspersonerna i stället föreställde sig en beröring av armen ökade aktiviteten i känselbarken (Davidson & Schwartz, 1977).
Liknande resultat kommer från blodflödesmätningar i hjärnan. Uppgifter som kräver visualisering tycks leda till ökad ämnesomsättning i hjärnans synbark. Goldenberg och medarbetare jämförde den regionala aktiviteten (mätt som blodflöde) i olika delar av hjärnan när försökspersoner fick besvara frågor som kan antas utlösa försök till visualisering ("Är det gröna på granen mörkare än det gröna hos gräset?") med sådana som inte gör det ("Är det kategoriska imperativet en gammaldags grammatisk form?"). Frågor av det första slaget ledde till en kraftigare ökning av aktiviteten i den primära synbarken (Farah, 1988)). En lång rad senare studier har visat att den primära synbarken aktiveras kraftigt när försökspersoner ombeds föreställa sig något eller återkalla en synbild i minnet (Le Bihan et al., 1993; Kosslyn et al., 1993).
Det bör i detta sammanhang också understrykas att den primära
synbarken, som tar emot signaler direkt från ögats näthinna,
faktiskt får nästa lika mycket inflöde från andra
delar av hjärnan. Fastän funktionen hos detta inflöde skulle
kunna vara att modifiera synintryck eller reglera en selektiv uppmärksamhet,
ligger det också nära till hands att tänka sig att det
är till för att aktivera synminnen och visuella fantasier.
Antecipation
Normal perceptuell aktivitet utlöses av stimulering av sinnesorganen, vilka i sin tur skickar signaler till de sensoriska områdena i hjärnbarken. Varifrån kommer då de signaler som utlöser perceptuell aktivitet i frånvaro av yttre stimuli?
En mekanism skulle kunna vara en association mellan olika stimuli. Det
finns experimentella belägg för att däggdjur kan lära
sig sådana associationer. Ett intressant exempel är s.k. sensorisk
prekonditionering. I ett klassiskt experiment presenterade Brogden först
upprepade gånger ett ljud och en ljussignal för en hund. Inget
av dessa stimuli framkallade någon synlig respons, så han kunde
inte i detta skede se om hunden hade lärt sig något. I ett andra
steg presenterades ljuset tillsammans med en elektrisk stöt mot tassen,
vilket utlöste en reflexmässig lyftning av foten. Så småningom
lyfte hunden tassen redan vid presentation av ljussignalen. När Brogden
nu presenterade ljudet för hunden, kom den också att lyfta på
tassen, trots att ljudet aldrig parats med någon stimulering av tassen
(Brogden, 1939). Tydligen hade hunden under den första experimentfasen
lärt sig associera ljud- och ljussignalerna. Det är inte känt
hur detta går till, men det är ingen långsökt hypotes
att ljudet, när det paras med ljussignalen, aktiverar någon
del av hjärnans synområden. En sådan mekanism skulle kunna
förklara att hörandet av ett ord kan leda till att man "ser"
det som ordet symboliserar.
 |
| Figur 2 |
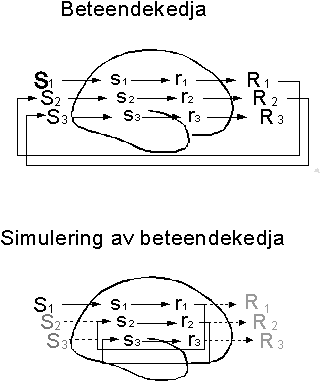
En alternativ mekanism, kanske mera grundläggande, är vad vi här kan kalla antecipation. Betrakta det enkla reaktionsschemat i figur 2, där en individ reagerar på en situation eller ett stimulus S med en viss respons R. (Denna terminologi missuppfattas ibland så att man tror att ett stimulus-responssamband måste stå för något enkelt och okomplicerat. I verkligheten är naturligtvis de flesta stimuli ytterst komplexa och innefattar många olika aspekter av den situation en organism befinner sig i. Även en respons kan vara komplex och bestå av många delar.) Många av dessa stimulus-responskopplingar har förvärvats genom enkla betingningsmekanismer. Mellan stimulus och respons ligger en rad komplexa händelser i hjärnans nervceller men vi förenklar denna process genom att säga att stimuleringen S orsakar en serie händelser s i hjärnans sensoriska delar vilka i sin tur leder till en serie händelser de motoriska delarna som vi kallar r vilka till sist leder till en fullbordad respons R. Vi kan tänka på s som aktivitet i de sensoriska bansystemen och i de delar av hjärnbarken som brukar betecknas som sensoriska, dvs nack-, tinning- och hjässloberna. Vi kan tänka på r som "förberedande" eller "incipient" beteende i frontallobernas premotoriska områden. Gränsdragningen mellan sensoriska och motoriska händelser är i hög grad godtycklig men har ingen betydelse för resonemanget här.
De sensoriska konsekvenserna av vårt beteende är ofta förutsägbara och mycket talar för att många organismer i en viss bemärkelse faktiskt kan förutsäga dem. Detta kan ta formen av rörelser för att undvika olika former av hinder i miljön. Ett djur som rör sig framåt i terrängen kan undvika att stöta ihop med en sten genom att hoppa över den. Sådant beteende skulle kunna beskrivas som en "antecipation" av en förväntad kollision. Detta kunde tyckas vara en onödig mystifiering av en process som kan beskrivas enkelt i termer av stimuli och responser. Stenen är ett stimulus och djuret har lärt sig att vid åsynen av ett sådant föremål hoppa över det och vi behöver inte alls inte postulera en ytterligare process för att förklara det. Det finns emellertid skäl att anta att det faktiskt existerar en substantiell form av antecipation.
Beteenden har alltid sensoriska konsekvenser. När en organism i
en viss situation utför ett beteende, kommer detta att förändra
situationen och generera nya stimuli, vilka orsakar ett nytt beteende och
så vidare (figur 3). Om det finns en något så när
regelbunden koppling mellan en viss en viss respons och dess sensoriska
konsekvenser, kommer det också att finnas en korrelation mellan motsvarande
interna händelser. Om alltså utförandet av responsen R
brukar följas av ett stimulus S, kommer utförandet av
den "förberedande" responsen r att regelbundet åtföljas
av en intern perceptuell aktivitet s. Den antecipationsmekanism
vi antar består i en förmåga att under tidigare stadier
av en respons utlösa en sensorisk aktivitet som liknar den som normalt
skulle utlösas av den fullbordade responsen. Det finns inget direkt
belägg för en sådan mekanism, men dess existens ter sig
ändå sannolik. För det första finns det rikligt med
förbindelser från frontalloben till alla sensoriska associationsområden
i hjärnan och från dessa till de primära sensoriska områdena.
För det andra finns det indirekta experimentella data som tyder på
att simulering av beteende kan aktivera sensoriska områden. I flera
studier av simulerade handlingar har man kunnat konstatera en samtidig
ökning av aktiviteten i sensoriska områden (Parsons et al.,
1995).
 |
| Figur 3 |
Vi antar således att den förberedande aktiviteten r i figur 3 efter hand automatiskt utlöser s. Vi antar också att denna internt utlösta sensoriska aktivitet är tillräckligt verklighetstrogen för att kunna utlösa en ny respons. Vilket värde skulle en sådan associationsmekanism ha?
Antag att vi har en organism som utför responsen R1 i situationen S1 och att detta resulterar i en ny situation S2. S2 är potentiellt farlig och organismen har lärt sig, att i närvaro av S2,utföra den defensiva responsen R2 (figur 3). Om denna händelsekedja inträffade ofta, skulle det vara värdefullt för organismen om det förutsägbara S2kunde simuleras som reaktion på den "incipienta" eller "förberedande" beteendet r1, som föregår R1, d.v.s. om r1 kunde utlösa s2 direkt, utan omvägen över öppet beteende och dess konsekvenser (figur 3B). Om s2, som vi nu antar, är tillräckligt lik den aktivitet som uppkommer när S2 inträffar, kommer s2 att utlösa r2 och den defensiva responsen R2 snabbare, i bästa fall innan S2 .
Låt oss översätta detta abstrakta resonemang till ett konkret exempel: Varje gång organismen ser ett hinder och går rakt emot detta, kolliderar det med hindret och tvingas till en undvikandemanöver. Efter att ha råkat ut för detta ett antal gånger, kommer förberedelsen för beteendet "att gå emot ett hinder" att utlösa en simulerad perception av kollisionen, vilket utlöser undvikandemanövern redan innan kollisionen inträffar.
Det är på intet sätt självklart att en snabb defensiv reaktion måste utlösas av just en simulerad perception. Organismen skulle också kunna lära sig att utföra den defensivareaktionen som svar på åsynen av hindret. En sådan mekanism existerar faktiskt i form av s.k. klassisk betingning och lärs in i lillhjärnan utan att sensoriska hjärnbarksområden behöver vara inkopplade (Hesslow & Yeo, 2001).
Det har emellertid stora fördelar att lära sig att inte bara utföra en defensiv reaktion i närvaro av vissa stimuli utan att faktiskt också "se" den farliga situationen. Skälet är informationsekonomiskt. Det kan finnas flera reaktioner som bör utföras i en viss situation. En antilopkalv som får syn på en leopard behöver springa fort i motsatt riktning, men den bör också tillkalla sin moder och dessutom söka i omgivningen efter flyktvägar. Antag vidare att det finns något stimulus som förvarnar om den farliga situationen, kanske ett karakteristiskt ljud som leoparden orsakar. Det vore naturligtvis värdefullt för kalven om också ljudet utlöste alla de defensiva reaktionerna. Om nu kalven vore tvungen att lära sig att associera leopardens ljud med var och en av de defensiva reaktionerna, skulle den behöva lära sig ett stort antal associationer: att vid ljudet av en leopard springa i motsatt riktning, att vid ljudet av en leopard ropa på mamma osv. Detta skulle kräva en tidsödande och framför allt farlig träning. En effektivare lösning vore att låta leopardljudet utlösa en simulering av åsynen av en leopard, vilket redan utlöser alla de önskade reaktionerna. Det räcker således med en association.
En antecipationsmekanism av detta slag kan också förklara
vissa experimentella resultat som ofta tolkats som bevis för existensen
av någon sorts mentala kartor ("cognitive maps") och mentala representationer
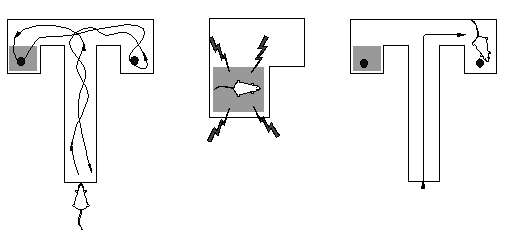
av mål för beteenden. I ett klassiskt experiment lät Tolman
en råtta undersöka en T-labyrint med en mörk kammare i
slutet av den vänstra armen och ljus kammare till höger (figur
5). Båda innehöll mat. Råttan placerades därefter
i en mörk kammare och utsattes för obehaglig stimulering i form
av elektriska stötar mot tassarna. När råttan senare placerades
i T-labyrinten gick den inte till den mörka kammaren, trots att den
aldrig bestraffats för detta beteende. Det är som om den
hade en bild av labyrinten och "visste" vad som fanns i slutet av den vänstra
armen (Tolman & Gleitman, 1949).
 |
| Figur 4 |
En enklare tolkning av detta experiment är att om råttan,
när den når T-korsningen, förbereder sig att gå till
vänster, kommer att utlösa en perception av den mörka kammaren
vilket, p.g.a. en tidigare betingning, utlöser ångest. Detta
undertrycker fullföljandet av den påbörjade responsen.
I en ganska oproblematisk och konkret mening kan man säga att råttan
"förutser" konsekvensen av att gå till vänster.
Simulering av interaktion med omvärlden
När väl antecipationsmekanismen finns på plats, är
det ingenting som hindrar att långa kedjor av simulerade responser
och perceptioner kan uppkomma. En simulerad handling i frontalloben genererar
en simulerad perception av handlingens sannolika konsekvenser i hjärnbarkens
sensoriska regioner. Denna perceptuella aktivitet fungerar som stimulus
för en ny handling (figur 5). En sådan kedja av simulerade stimuli
och responser kan i princip fortgå hur länge som helst eller
tills den bryts av någon extern faktor.
 |
| Figur 5 |
Antag, för att ta ett konkret exempel, att jag sitter i mitt arbetsrum och vrider huvudet åt vänster eller höger. Varje rörelse utlöser en specifik visuell stimulering. När jag vänder mig åt vänster ser jag ett fönster; när jag tittar åt höger ser jag dörren. Om någon ropar mitt namn från höger, utlöses aktivitet i min hörselbark; detta leder i sin tur till aktivitet i premotoriska områden i frontalloben, vilket leder till att den primära motoriska barken får mig att vrida huvudet åt höger; jag får nu en ny visuell stimulering från dörren som får mig att resa mig och öppna dörren för den person som står utanför. Säg nu att jag i stället "tänker" att jag hör en röst från dörren, dvs simulerar hörandet i min hörselbark. Detta leder till att min frontallob börjar förbereda en huvudrörelse, men redan innan denna kommer igång utlöses en simulerad perception av dörren. Låt mig illustrera med ytterligare ett par exempel.
Schackspel
Låt oss tänka oss en person som prövar ut en ny schackstrategi. Han betraktar konfigurationen av pjäserna på schackbrädet och prövar ett visst drag. Detta skapar en ny konfiguration, som omedelbart utlöser ett motdrag, som leder till en katastrofal tredje konfiguration och en förödmjukande förlust. I ett verkligt spel finns det ingen möjlighet att fysiskt pröva olika spelstrategier och en spelare får därför nöja sig med att göra det näst bästa, nämligen att simulera dem. När han betraktar den ursprungliga pjäsuppställningen kommer de premotoriska områdena i hans frontallob att aktiveras som förberedelse för ett drag, men utflödet hämmas så att draget aldrig fullbordas. Frontallobsaktiviteten är emellertid tillräcklig för att generera den visuella aktivitet som skulle ha uppkommit om draget hade fullbordats och tillåtits förändra pjäsuppställningen. Denna simulerade visuella aktivitet är tillräcklig för att utlösa ett nytt drag som leder till en ny konfiguration på schackbrädet. Naturligtvis kan bara en tränad schackspelare göra detta, därför att bara en tränad schackspelare har övat upp de associationer som är nödvändiga för att ett visst drag skall generera en relevant perception. Förmågan att vidmakthålla en tillräckligt väl simulerad perception, för att denna skall utlösa ett adekvat nästa drag, är säkert också beroende av träning.
Inre tal
Trots att en viktig del av vår inre värld består i verbalt tänkande, är det inte nödvändigt att vara medveten för att tala. Om man snabbt skall svara på en enkel fråga, vad klockan är, säg, eller vad jag heter, kommer svaret normalt automatiskt och mycket snabbt och behöver inte föregås av några medvetna tankar. Men om förberedelsen för ett verbalt yttrande kan generera aktivitet i hörselbarken, kan man höra sitt eget tal innan det har hunnit fullbordas (eller utan att det fullbordas). Det borde därför vara möjligt att simulera en konversation med sig själv (Hesslow, 1994).
En annan konsekvens av detta är att talaren kan sägas "höra" sitt yttrande två gånger, först som en internt genererad perception och strax därefter som en verklig perception via hörseln av det fullbordade yttrandet. Det är ingen långsökt hypotes att detta kan bidra till föreställningen att talet föregås av en tanke. På sätt och vis är detta korrekt, om vi nu vill kalla den simulerade perceptionen en tanke, men det är förstås ett misstag att tro att denna tanke är en orsak till det färdiga yttrandet. I en viss mening är det tvärtom så att (förberedelsen för) yttrandet orsakar "tanken". När vi anser oss vara medvetna om vad vi säger, är det kanske just denna antecipation av vad vi strax skall säga som är den avgörande mekanismen.
Man får förutsätta att simulerade perceptioner är
mindre effektiva än sådana som utlöses från sinnesorganen,
men om de bara är tillräckliga för att utlösa adekvata
beteendesimuleringar, behöver en sådan kedja inte bli svagare
bara därför att den blir längre. Däremot ökar
naturligtvis sannolikheten för att en dold respons skall uppkomma
för vilken ingen inlärd perception kan utlösas. Detta bryter
naturligtvis kedjan.
Abstrakt beteende
Vi har tidigare talat om simulerade handlingar som i princip samma sak som ofullbordade handlingar eller handlingar, där de förberedande processerna i frontalloberna äger rum, som vid fullbordade handlingar, men där utflödet från den primära motoriska barken är undertryckt. Man kan emellertid tänka sig, att denna undertryckning äger rum på ett tidigare stadium, vilket skulle kunna ge upphov till vad vi kan kalla abstrakta handlingar.
Låt oss säga att en person skall rita en triangel. Någonstans i frontalloberna genereras en signal för detta som ger upphov till mer detaljerade signaler längre bak för att ta fram en penna och utföra själva ritandet och därefter signaler för hur den ena handen och fingrarna skall röra sig för att resultatet skall bli rimligt likt en triangel. Allt detta kan ske utan att vi faktiskt utför någon rörelse. På varje nivå genereras sensoriska konsekvenser av handlingen, först "åsynen" av en triangel, därefter "ser" vi hur handen fattar en penna och hur den ritar och vi "känner" handens och fingrarnas rörelser. Men om man kan undertrycka effektuerandet av denna handling precis innan de motoriska nervcellerna i den primära motorbarken aktiveras, borde man i princip kunna undertrycka den långt tidigare, redan innan de signaler uppkommer som genererar specifika hand- och fingerrörelser. Kanske det blir möjligt att med träning utföra bara det allra första stadiet, signalen att en triangel skall skapas. En sådan handling blir då relativt frikopplad från den motoriska fortsättningen och det blir kanske något vilseledande att beskriva den som en simulering av beteende. Härav benämningen "abstrakt".
Även när det gäller perceptuell aktivitet skulle man kunna tänka sig en mer eller mindre långtgående frikoppling från externa stimuli. När vi föreställer oss ett konkret föremål eller en konkret händelse aktiveras, som tidigare nämnts, den primära synbarken. Vi kan också föreställa oss abstrakta egenskaper utan att samtidigt tänka på dessa som egenskaper hos ett konkret objekt. Man kan tänka på en triangel utan att tänka på något specifikt triangulärt föremål eller på en bild av triangel på ett papper. Man kan se blått utan att se ett blåfärgat föremål. Det är en spekulation, men alls inte långsökt, att man vid sådana tillfällen aktiverar de nervceller i visuella associationsområden som normalt aktiveras av abstrakta egenskaper hos det man ser. En sådan process skulle kunna ligga bakom uppkomsten av faktaminnen ur minnen av händelser.
En relativ frikoppling av dolda eller simulerade handlingar från
fysiska handlingar och av fantasier från faktisk perception av yttervärlden
torde medföra en avsevärd ökning av vår kognitiva
förmåga utan att egentligen förutsätta någon
ny fysiologisk mekanism. Kanske en ökad förmåga att göra
denna frikoppling är en del av förklaringen till skillnaden i
kognitiv förmåga mellan människor och andra primater.
Initiering och utvärdering av simulering
Det finns ingenting i de ovan beskrivna simuleringsmekanismerna, som inte skulle kunna implementeras i en enkel organism eller en mekanisk konstruktion. Antagligen är det frestande för många att tänka sig, att det måste finnas ett "jag" som avsiktligt sätter igång simuleringskedjan och utvärderar dess resultat, en autonom aktör som står utanför kedjan av stimuli och responser. Det ligger utanför syftet med denna uppsats att i detalj beskriva hur en simulering kan initieras och utvärderas. Det är dock nödvändigt att visa att det, åtminstone i princip, är möjligt att redogöra för detta utan att smuggla in ett spöke i maskineriet.
Initiering av beteendesimulering sker normalt genom det vi kallat antecipation. De flesta av våra handlingar genererar automatiskt svaga upplevelser av sina normala konsekvenser. Detta innebär att det första steget i en simulering initieras av samma situation som initierar en handling och en möjlig mekanism för initiering av en simuleringskedja är att detta första steg helt enkelt automatiskt följs av andra. Redan när jag tar det första steget mot kylskåpet, är jag "i tankarna" på väg att öppna det, ta ut en bit salami och göra mig en smörgås. Mycket, kanske det mesta av vårt medvetna "tänkande" består i att vi fullföljer långsiktiga konsekvenser, inklusive adekvata handlingar och förväntade upplevelser, av handlingar vi redan har satt igång.
I sådana fall är en simulering egentligen inte en handling i sig, utförd i avsikt att utvärdera konsekvenserna av ett möjligt handlingsalternativ. Vi kan emellertid också lära oss att genomföra simuleringar som en särskild form av beteende. I situationer där flera alterntiva handlingar konkurrerar men ingen automatiskt "vinner", kan vi lära oss att systematiskt gå igenom alternativen och fullfölja konsekvenserna av vart och ett.
Också utvärderingen av en simulering kan tyckas förutsätta
en autonom agent, som utför utvärderingen, men detta är
faktiskt inte nödvändigt. Vissa konsekvenser av handlingar, även
om de bara är inbillade, utlöser emotionella reaktioner, behagliga
eller obehagliga. En möjlig effekt av detta skulle kunna vara att
en enkel mekanism för operant betingning förstärker eller
försvagar den simulerade handling som har den aktuella konsekvensen.
Eftersom den simulerade handlingen bara är en svagare form av den
fullbordade, kommer även den senare att förstärkas eller
försvagas. En annan möjlighet är att den emotionella konsekvensen
fungerar som ett (diskriminativt) stimulus. Vi har helt enkelt lärt
oss att det är bra att utföra sådana handlingar vars simuleringar
har behagliga konsekvenser och undvika dem som skapar ångest. (För
en mycket läsvärd diskussion av detta problem, se Damasio, 1994).
Oavsett vilken den exakta mekanismen är, står det klart att
vi inte behöver förutsätta något jag eller någon
annan autonom agent för att förklara vare sig initiering eller
utvärdering av en simulerad kedja av handlingar och perceptioner.
Fördelar med simuleringshypotesen
Utöver den empiriska evidens som relaterats ovan och utöver
dess förmåga att besvara de frågor om medvetandet och
den inre världen som inledde denna uppsats (se nedan), vill jag peka
på tre attraktiva egenskaper hos simuleringshypotesen.
Den kräver inga avancerade teoretiska antaganden.
När behaviorismen föll ur modet på 1960-talet och den
sk "kognitiva revolutionen" svepte genom psykologin, minskade kravet på
teoretisk försiktighet och det blev legitimt att göra synnerligen
långtgående antaganden om mentala processer och storheter.
Den kognitivistiska litteraturen flödar över av begrepp som "informationsbearbetning",
"representationer", "kognitiva kartor", "mentala modeller av yttervärlden"
och mycket annat med bräckligt empirisk underlag och svag fysiologisk
förankring. Simuleringshypotesen kanske inte kan förklara alla
de fenomen som dessa begrepp har införts för att hantera, men
den förklarar många av dem, trots att den helt och hållet
är formulerad i fysiologiska och anatomiska termer. Denna förankring
i empirisk biologi och frånvaron av hypotetiska kognitiva storheter
är en avgjord styrka.
Den förutsätter inga evolutionära språng.
Det är inte mer än 1.24% av vår arvsmassa som skiljer en människa från en schimpans och det finns således inte mycket utrymme för några stora evolutionära språng. Om medvetande och kognitiva processer inte är sekundära till de grundläggande sensoriska och motoriska mekanismer som finns hos andra däggdjur, är det svårt att se att de skulle kunna uppkomma utan ganska omfattande förändringar av hjärnan. Precis som man skulle förvänta sig mot bakgrund av de ringa genetiska skillnaderna, har det emellertid inte tillkommit några områden i människans hjärna som inte också finns hos alla andra däggdjur. Alla de stora strukturerna, hippocampus, cerebellum, basala ganglier osv, finns hos alla däggdjur och ser i stort sett likadana ut. Den enda tydliga skillnaden är kvantitativ. Storhjärnans bark och vissa delar av övriga strukturer är större, men grundkonstruktionen är densamma. Det måste vara en styrka hos simuleringshypotesen att den förklarar hur kognitiva funktioner har uppkommit med hjälp av strukturer och mekanismer som finns hos alla däggdjur.
Ytterligare några ord om djurs kognitiva förmåga är befogade. En viktig komponent i den syn på medvetande och tänkande, som jag har redogjort för ovan, är att det inte finns någon principiell skillnad mellan människor och andra däggdjur, i synnerhet inte mellan människor och andra primater. Naturligtvis kan effektiviteten eller hastigheten i de associativa mekanismerna skilja sig och kanske är likheten mellan perceptuell simulering och perception större hos människan än hos andra arter, men det är här fråga om gradskillnader, inte om principiella skillnader. De variationer, som finns mellan olika arter i kognitiv förmåga, framstår från denna utgångspunkt som sekundära och perifera och säger mera om olika arters levnadsförhållanden än om deras kognitiva basutrustning. Liksom vi inte gör något stort nummer av att svenskar har svårt att förstå kinesisk grammatik har det inte någon stor teoretisk betydelse att apor inte bygger hus eller skriver böcker.
Vissa forskare hävdar emellertid att skillnaderna mellan människor och andra djur är djupgående och av fundamental betydelse för förståelsen av människans kognitiva förmågor. Povinelli (2000) hävdar t.ex. att schimpanser är oförmögna att "förstå" även mycket enkla fysikaliska principer. Om en schimpans behöver fösa ett föremål över en yta som innehåller en fördjupning, förstår den inte att föremålet kommer att ramla ner i fördjupningen. Staplar den lådor på varandra förstår inte apan att stapeln rasar om man drar ut den understa lådan. Inte ens efter omfattande träning kan schimpanser klara sådana uppgifter.
Den "träning" som kan komma ifråga för en schimpans
kan dock inte jämföras med den träning som ett barn erhåller.
Mänskliga barn ägnar flera timmar om dagen i många år
åt mekaniska lekar. Därvid övar de upp sin förmåga
att hantera enkla föremål och tränar in de sensoriska konsekvenserna
av olika manipulationer. Det är inlärningen av dessa associationer
som gör det möjligt för dem att senare antecipera och undvika
konsekvenserna av att fösa föremål över fördjupningar
och dra undan den understa lådan i en stapel. Det finns direkta empiriska
belägg för att människor drar slutsatser av det slag som
Povinelli studerade just genom att simulera handlingar (Schwartz &
Black, 1999).
Den förklarar sambandet mellan motorik och kognition.
Skador på lillhjärnan och de basala ganglierna ger dramatiska
och mycket karakteristiska motoriska symptom. Förändringar i
lillhjärnan ger t.ex. en oförmåga att koordinera olika
muskler. Parkinsons sjukdom, som orsakas av förändringar i de
basala ganglierna, kännetecknas bl.a. av en svårighet att initiera
rörelser. Det har lett till ganska stor förvåning att dessa
delar av hjärnan också tycks vara inblandade i kognitiva funktioner.
När man med exempelvis blodflödesmätningar studerar vilka
delar av hjärnan som engageras vid olika kognitiva uppgifter finner
man ofta, att såväl lillhjärnan som de basala ganglierna
aktiveras (Decety et al., 1990). I överensstämmelse härmed
har man också funnit i kliniska studier från senare år
att patienter med skador i hjärnans motoriska delar uppvisar kognitiva
symptom som motsvarar de motoriska. Hos patienter med Parkinsons sjukdom
är t.ex. tänkta rörelser hämmade på samma sätt
som utförda rörelser och patienter med lillhjärneskador
har störningar i amplituden hos kognitiva aktiviteter som påminner
om de typiska motoriska symtomen (Dominey et al., 1995; Schmahmann,
1997).
Medvetandets problem
Hur uppkommer den inre världen? Svaret på denna fråga är nu banalt. Den inre världen uppkommer genom att hjärnan själv kan generera sitt eget inflöde när den simulerar handlingar och deras sensoriska konsekvenser. Vi kan "se", "höra" och "känna" konsekvenserna av det vi gör trots att dessa (ännu) inte existerar. Observera att denna slutsats är oberoende av hur perceptionen faktiskt går till. Vi behöver t.ex. inte ta ställning till om en fantasiföreställning har karaktären av en bild eller inte. Om hjärnan kan generera sina egna stimuli, måste en inre värld uppkomma oavsett hur dessa stimuli faktiskt bearbetas. Det är dock fördel med det synsätt jag försökt förklara och försvara i denna uppsats, att det innebär ett minimum av ontologiska förutsättningar. Den är således helt förenligt med ett strikt behavioristiskt synsätt. Vi behöver inte anta några mentala storheter, inte ens interna "bilder" eller "representationer".
Genom att simuleringen innebär att de delar av hjärnan aktiveras som normalt utför beteenden och bearbetar sensoriska intryck, så blir det självklart att det är hjärnans motoriska och sensoriska delar som ligger bakom även det medvetna tänkandet och det finns inget behov av några särskilda strukturer i hjärnan för medvetande, tänkande eller problemlösande. Det bör inte heller förvåna oss att andra strukturer som traditionellt brukar betraktas som renodlat motoriska, som primära motorcortex, cerebellum och de basala ganglierna, aktiveras vid "kognitiva" funktioner. Dessa strukturer gör nämligen i hög grad samma saker vid simulerat som vid fullbordat beteende.
Vad är mentala objekt? Svaret är att vi inte behöver anta att tanken på ett visst objekt förutsätter existensen av ett motsvarande objekt. Patienten med en fantomsmärta i fingret har en perception av en skada, men vi behöver inte säga att det måste existera en "mental" eller fiktiv skada för att förstå hur detta är möjligt. Om jag manipulerar hastighetsmätaren i min bil så att den får en signal som gör att den visar 100 km/t när bilen står stilla, så är det ur displayens "synvinkel" ingen skillnad mellan att registrera en verklig hastighet av 100 km/t och en "simulerad". Vi kan förstå detta utan att anta existensen av en mental hastighet.
Vad är en tanke? Ett enkelt svar är att en tanke kan beskrivas som en förberedelse för handling åtföljd av perceptionen av handlingens mest sannolika konsekvenser, om den hade fullbordats. Tänkande är kedjor av sådana simulerade handlingar och handlingskonsekvenser.
Funktionsproblemet. Förmågan att simulera handlingar och deras konsekvenser har ett uppenbart värde för oss. Det är så vi prövar och utvärderar olika handlingsalterntiv utan att behöva ta risker och med mindre ansträngning och tid. Eftersom processen generar stimuli som liknar dem, som vid perception genereras av yttervärlden, kommer en inre värld, som påminner om yttervärlden, att bli en oundviklig konsekvens av denna förmåga.
Vilken sorts varelser kan ha en inre värld? Alla organismer,
såväl biologiska som artificiella, som internt kan generera
stimuli, t.ex. genom att simulera beteende, kommer att ha en inre värld,
låt vara att denna kan vara ganska torftig jämfört den
mänskliga motsvarigheten. Det förefaller mycket sannolikt att
alla däggdjur, och kanske också många lägre ryggradsdjur,
har förmågan att antecipera konsekvenserna av sina handlingar.
I så fall har de en inre värld. Rizolattis experiment talar
för att apor kan simulera handlingar och anger en tänkbar väg
att undersöka om också andra däggdjur kan göra det.
Det är naturligtvis också möjligt att konstruera en robot
efter sådana principer. Försök att simulera en sådan
robot har redan utförts (Jirenhed et al., 2001).
Slutord
Säkert kommer många läsare att tycka, att det finns
många frågor om medvetandet, som de inte fått besvarade.
Det har dock inte varit min ambition att formulera en hypotes om "medvetandet"
som besvarar alla sådana frågor. Som jag inledningsvis förklarat
tror jag inte att något sådant är möjligt. Det jag
har försökt göra troligt är, att medvetandeproblemet
består av många frågor, varav vissa kan ges tillfredsställande
svar redan nu, bland annat genom en mekanism för simulering. Det förefaller
troligt att fysiologiska mekanismer kommer att belysa även de problem
som återstår.
Not
De idéer som presenteras i denna uppsats är ett förtydligande
av tankar som tidigare presenterats i Hesslow (1994) och Hesslow (1996).
De har i hög grad formats under samtal med psykologen Håkan
Ornander med vilken jag diskuterat medvetandet av och till under ett antal
år. Vi har i vår tur båda inspirerats av idéer
som formulerats i den behavioristiska traditionen, fa. av B.F. Skinner
(1953; 1957; 1974). Under skrivandet har jag upptäckt att närbesläktade
idéer har formulerats av många andra som Alexander Bain (1868)och
William James (1890). Även några samtida författare har
fört fram liknande tankegångar, t.ex. Donahoe & Palmer (1994).
Jag vill tacka Fredrik Bengtsson, Dan-Anders Jirenhed och Magnus Larsson
för värdefulla diskussioner och synpunkter på tidigare
versioner av denna uppsats. Arbetet har stötts av Medicinska forskningsrådet
(projekt 09899).
Bain, A. (1868). The Senses and the Intellect, 3 ed. Longmans, Green & Co, London.
Bisiach, E. & Luzzatti, C. (1978). Unilateral neglect of representational space. Cortex 14, 129-133.
Bonnet, M., Decety, J., Requin, J., & Jeannerod, M. (1997). Mental simulation of an action modulates the excitability of spinal reflex pathways in man. Cognitive Brain Research 5 , 221-228.
Brogden, W. J. (1939). Sensory preconditioning. Journal of Experimental Psychology 25, 323-332.
Damasio, A. R. (1994). Descartes' Error: Emotion, Reason and the Human Brain G.P. Putnam's Sons, New York.
Davidson, R. J. & Schwartz, G. E. (1977). Brain mechanisms subserving self-generated imagery: Electrophysiological specificity and patterning. Psychophysiology 14, 598-601.
Decety, J., Jeannerod, M., & Prablanc, C. (1989). The timing of mentally represented actions. Behavioural Brain Research 34, 35-42.
Decety, J., Sjöholm, H., Ryding, E., Stenberg, G., & Ingvar, D. H. (1990). The cerebellum participates in cognitive activity: Tomographic measurements of regional cerebral blood flow. Brain Research 535, 313-317.
Deiber, M. P., Ibanez, V., Honda, M. S. N., Raman, R., & Hallett, M. (1998). Cerebral processes related to visuomotor imagery and generation of simple finger movement studied with positron emission tomography. Neuroimage 7, 73-85.
Di Pellegrino, G., Fadiga, L., Fogassi, L., Gallese, V., & Rizzolatti, G. (1992). Understanding motor events: A neurophysiological study. Experimental Brain Research 91, 176-180.
Dominey, P. F., Decety, J., Broussolle, E., Chazot, G., & Jeannerod, M. (1995). Motor imagery of a lateralized sequential task is asymmetrically slowed in hemi-Parkinson patients. Neuropsychologia 33, 727-741.
Donahoe, J. W. & Palmer, D. C. (1994). Learning and Complex Behavior Allyn & Bacon, Needham Heights.
Farah, M. J. (1988). Is visual imagery really visual? Overlooked evidence from neuropsychology. Psychological Review 95, 307-317.
Hesslow G. & Yeo, C. H. (2001). The anatomy and physiology of classical conditioning. In Classical Conditioning: A Source Book, ed. Moore, J. E., Springer, Berlin.
Hesslow, G. (1994). Will neuroscience explain consciousness? Journal of Theoretical Biology 171, 29-39.
Hesslow, G. (1996). Hjärnan och själen: kan fysiologin förklara den inre världen. In En bok om hjärnan, ed. Widén, L., Tiden, Stockholm.
Ingvar, D. H. & Philipsson, L. (1977). Distribution of the cerebral blood flow in the dominant hemisphere during motor ideation and motor performance. Annals of Neurology 2 , 230-237.
Jacobson, E. (1930). Electrical measurements of neuromuscular states during mental activities. I. Imagination of movement involving skeletal muscle. American Journal of Physiology 91, 567-608.
James, W. (1890). Principles of Psychology Macmillan (republished by Dover 1950).
Jeannerod, M. (1994). The representing brain: Neural correlates of motor intention and imagery. Behavioral and Brain Sciences 17, 187-245.
Jirenhed, D.-A., Hesslow, G., & Ziemke, T. (2001). Exploring internal simulation of perception in a mobile robot. Eurobot01.
Kosslyn, S. M. (1994). Image and Brain: The Resolution of the Imagery Debate MIT Press, Cambridge.
Kosslyn, S. M., Alpert, N. M., Thompson, W. L., Maljkovic, V., Weise, S. W., Chabris, C. F., Hamilton , S. E., Rauch, S. L., & Buonanno, F. S. (1993). Visual Mental Imagery Activates Topographically Organized Visual Cortex: PET Investigations. Journal of Cognitive Neuroscience5, 263-287.
Landauer, T. K. (1962). Rate of implicit speech. Perceptual and Motor Skills 15, 646.
Lashley, K. S. (1951). The problem of serial order in behavior. In Cerebral Mechanisms in Behavior, ed. Jeffress, L. A., pp. 112-146. Wiley, New York.
Le Bihan, D., Turner, R., Zeffiro, T. A., Cuénod, C. A., Jezzard, P., & Bonnerod, V. (1993). Activation of human primary visual cortex during visual recall: A megnetic resonance imaging study. Proceedings of the National Academy of Sciences 90, 11802-11805.
Monod, J. (1970). Chance and Necessity; An Essay on the Natural Philosophy of Modern Biology. (Le hasard et la nécessité : essai sur la philosophie naturelle de la biologie moderne) Seuil, Paris.
Parsons, L. M., Fox, P. T., Downs, J. H., Glass, T., Hirsch, T. B., Martin, C. C., Jerabek, P. A., & Lancaster, J. L. (1995). Use of implicit motor imagery for visual shape discrimination as revealed by PET. Nature375, 54-58.
Pinker, S. (1994). The Language Instinct Morrow, New York.
Povinelli, D. J. (2000). Folk Physics for Apes. The Chimpanzees Theory of How The World Works Oxford University Press, Oxford.
Pylyshyn, Z. W. (1984). Computation and Cognition MIT Press, Cambridge, MA.
Schmahmann, J. D. ed. (1997). The Cerebellum and Cognition Academic Press, New York.
Schwartz, D. L. & Black, T. (1999). Inferences through imagined actions. Knowing by simulated doing. Journal of Experimental Psychology [Hum Perc Perf] 25, 116-136.
Shepard, R. N. & Metzler, J. (1971). Mental rotation of three-dimensional objects. Science 171, 701-703.
Skinner, B. F. (1953). Science and Human behavior Macmillan, New York.
Skinner, B. F. (1957). Verbal Behavior Appleton-Century-Crofts, New York.
Skinner, B. F. (1974). About Behaviorism Knopf, New York.
Smith, S. M., Brown, H. O., Toman, J. E., & Goodman, L. S. (1947). The lack of cerebral effects of d-tubercurarine. Anesthesiology8, 1-14.
Sokolov, A. N. (1972). Inner Speech and Thought (translated from the Russian by G.T. Onischenko) Plenum Press, New York.
Tolman, E. C. & Gleitman, H. (1949). Studies in learning and motivation: I. Equal reinforcements in both end-boxes, followed by shock in one end-box. Journal of Experimental Psychology 39, 810-819.
Watson, J. B. (1930). Behaviorism, 2nd ed. Norton (Originally
published 1924), New York.